

 Home
Home

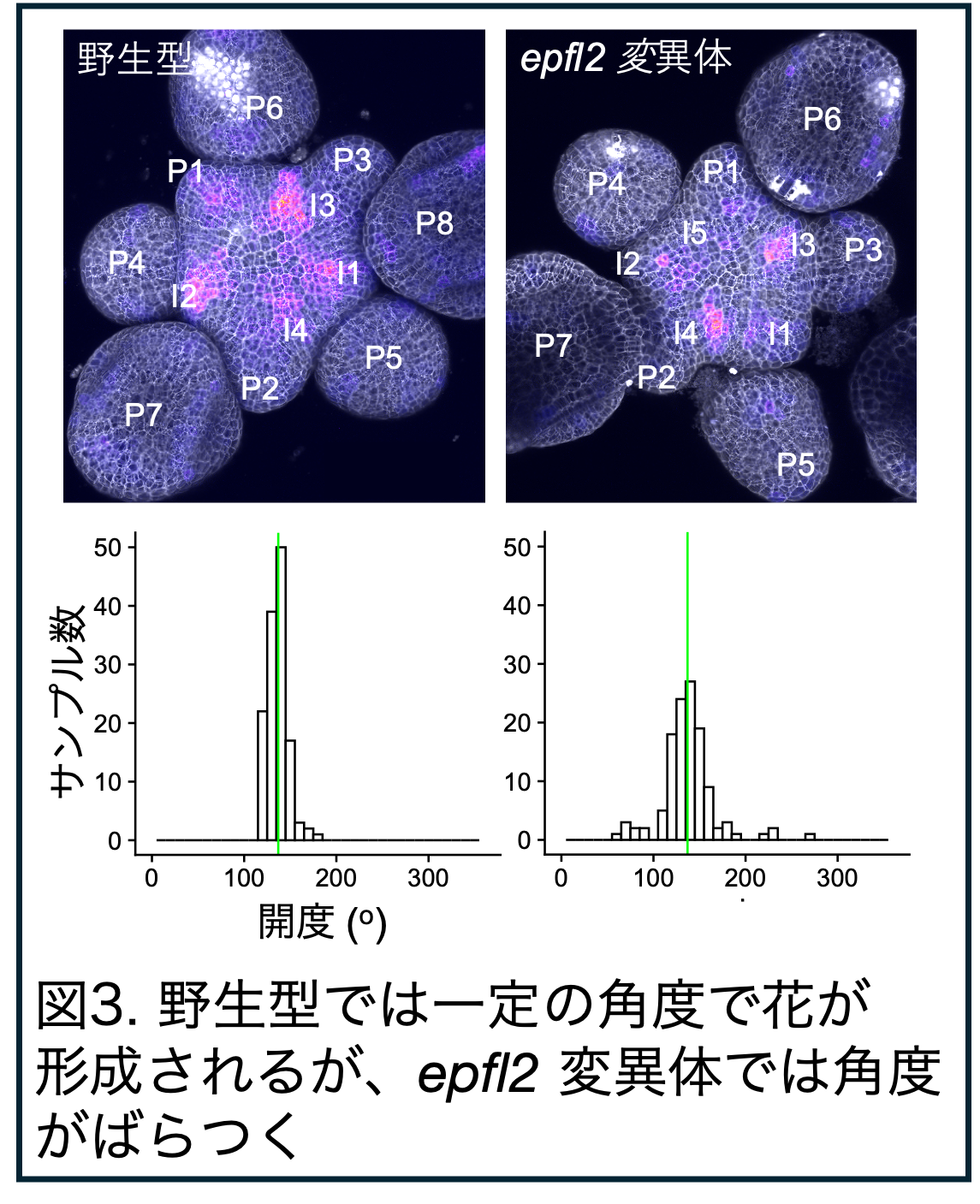
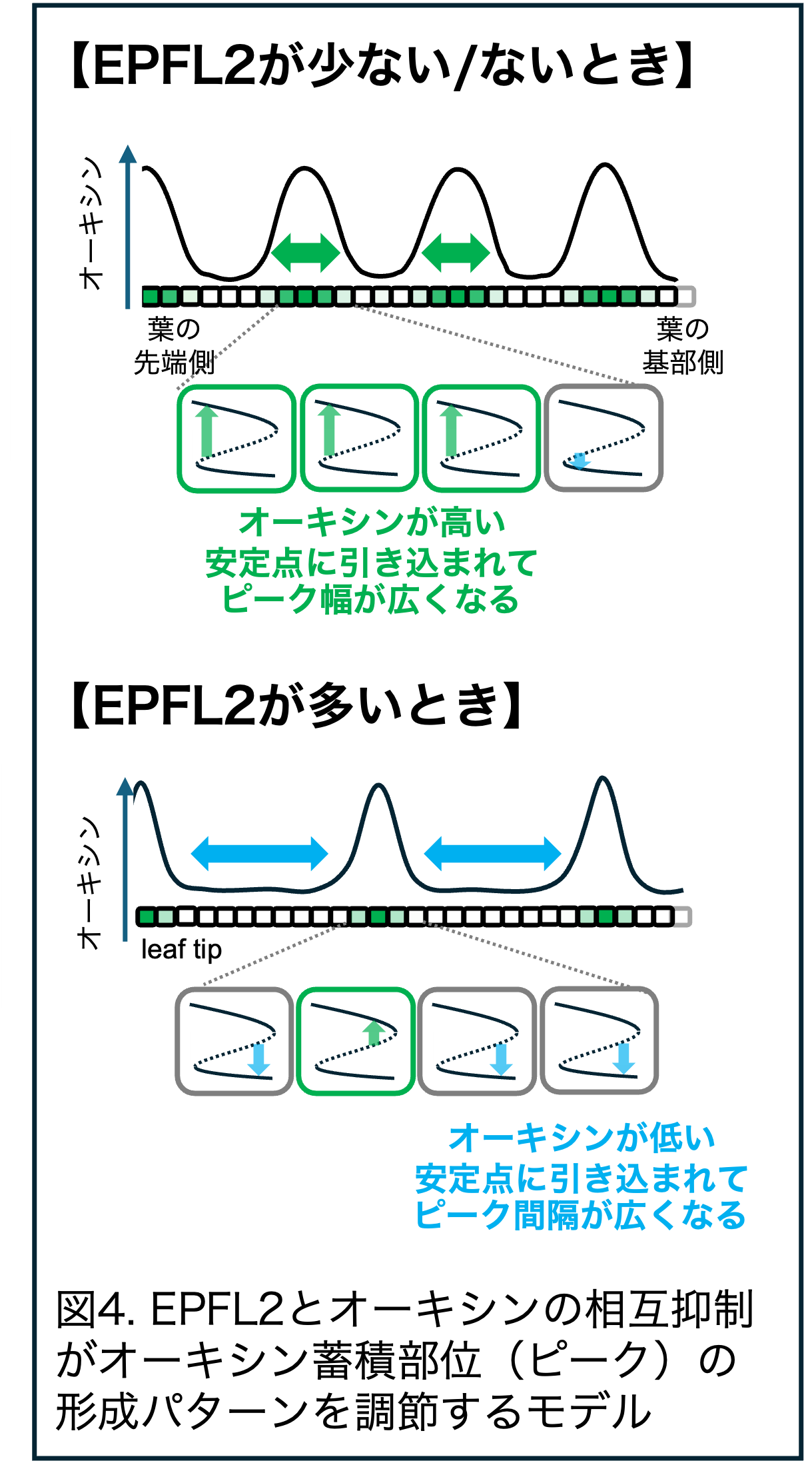
<研究に関すること>
奈良先端科学技術大学院大学 先端科学技術研究科 バイオサイエンス領域 植物再生学研究室
特任准教授 池内桃子
TEL:0743-72-5520 E-mail:momoko.ikeuchi@bs.naist.jp
研究室紹介ホームページ:https://bsw3.naist.jp/ikeuchi/
京都府立大学大学院 生命環境科学研究科
講師 爲重才覚
TEL:075-703-5444 E-mail:t-tamesige@kpu.ac.jp
研究室紹介ホームページ:https://sites.google.com/view/tameshigelab/
広島大学大学院統合生命科学研究科
教授 藤本仰一
TEL:082-424-7346 E-mail:kfjmt@hiroshima-u.ac.jp
<JST事業に関すること>
科学技術振興機構 創発的研究推進部 東出学信
TEL:03-5214-7276 E-mail:souhatsu-inquiry@jst.go.jp
<報道に関すること>
奈良先端科学技術大学院大学 企画総務課 渉外企画係
TEL:0743-72-5063/5112 FAX:0743-72-5011 E-mail:s-kikaku@ad.naist.jp
科学技術振興機構 広報課
TEL:03-5214-8404 FAX:03-5214-8432 E-mail:jstkoho@jst.go.jp
京都府立大学 企画・地域連携課 企画・地域連携係
TEL:075-703-5147 FAX:075-703-4979 E-mail:kikaku@kpu.ac.jp
名古屋大学 総務部広報課
TEL:052-558-9735 FAX:052-788-6272 E-mail:nu_research@t.mail.nagoya-u.ac.jp
広島大学広報室
E-mail:koho@office.hiroshima-u.ac.jp




