平成25年2月15日
国立大学法人東北大学
国立大学法人京都大学
国立大学法人広島大学
独立行政法人理化学研究所
公益財団法人高輝度光科学研究センター
X線自由電子レーザー (XFEL)を照射したタンパク質微結晶中の硫黄原子からの異常シグナルの検出に成功!
―XFELによるタンパク質分子の構造決定に向けた第一歩―
- タンパク質微結晶中の硫黄原子によるX線自由電子レーザー (XFEL)の異常散乱信号検出に成功
- 日本のXFEL施設 SACLAを用いたタンパク質微結晶構造解析法の発展に寄与
- XFELによる構造未知のタンパク質分子の構造決定への貢献が期待される
マックス・プランク研究所(Peter Gruss所長)、東北大学(里見進総長)、京都大学(松本紘総長)、広島大学(浅原利正学長)、理化学研究所(野依良治理事長)と高輝度光科学研究センター(白川哲久理事長)による合同研究チームは、X線自由電子レーザー(XFEL)*1をタンパク質分子の微結晶に照射して得られるX線回折*2において、タンパク質分子内に天然に含まれる硫黄原子による微弱な異常散乱*3信号を検出することに成功しました。本研究はマックス・プランク研究所Ilme Schlichting教授らのグループ、東北大学多元物質科学研究所上田潔教授のグループ、京都大学大学院理学研究科八尾誠教授のグループ、広島大学大学院理学研究科和田真一助教、理研放射光科学総合研究センターXFEL研究開発部門ビームライン研究開発グループ矢橋牧名グループディレクター及び高輝度光科学研究センター XFEL研究推進室 利用技術開発・整備チーム 登野健介チームリーダーのグループ等からなる合同研究チームによる成果です。
これまでに多くのタンパク質分子の構造がシンクロトロン放射光施設においてX線結晶構造解析法*4を用いて決定されてきましたが、結晶化が困難なために未だに構造が決定されていないタンパク質分子も多々あります。XFELの非常に強力なX線パルスを用いると非常に小さな結晶や結晶化していない試料からでもX線回折像を得ることができるため、これまで構造が決定できなかったタンパク質分子の構造が決定できると期待されています。しかし、XFELで得られるX線パルスのX線回折データから構造未知のタンパク質分子の構造を決定するためには、X線回折現象を表す構造因子の位相*5を決定する方法を確立しなければなりません。今回のタンパク質微結晶を用いた研究で検出された異常散乱信号を用いると位相を決定できるため、本研究はXFELを用いたタンパク質分子の構造決定に向けた第一歩に繋がると期待されます。
本研究の成果は、科学雑誌『Acta Crystallographica Section D』に近日中に掲載されます。
本研究は、合同研究チームの上田を代表とする文部科学省 X線自由電子レーザー利用推進研究課題、 理化学研究所SACLA利用装置提案課題、文部科学省 X線自由電子レーザー重点戦略研究課題の各事業、及び八尾を代表とする日本学術振興会二国間交流事業の一環として行われました。
米国のX線自由電子レーザー(XFEL)施設 LCLS*6に続いて、日本のXFEL施設 SACLA*7が完成し、我が国でも非常に強力かつパルス幅(X線の発光時間)の非常に短いパルスX線が利用できるようになりました。このXFELを利用すると、たとえば、化学変化において超高速で起こる個々の原子の動きのように、これまで見えなかった超極細、超高速なものが見えるようになると期待されています。その中でも、XFELを用いることによって大きな飛躍が期待されている分野がX線構造生物学です。これまで、シンクトロン放射光施設が世界中に次々に建設されるとともに、多くのタンパク質分子の構造がX線結晶構造解析法を用いて決定されてきました。一方で、結晶化が困難なために未だに構造が決定されていないタンパク質も多々あります。XFELの非常に強力なX線パルスを用いると非常に小さな結晶や結晶化していない試料からでもX線回折像が得られることから、これまで構造がわからなかったタンパク質の構造が決定できると期待されています。しかし、これを実現するためにはいくつかの障害を乗り越えなければなりません。特に問題となるのがX線回折現象を表す構造因子の位相の決定です。ごく最近、LCLSで初めてタンパク質の新規結晶構造が決定され話題になりましたが、その研究では、分子置換法と呼ばれる方法を用いて、すでに分かっている構造をもとにこの新規構造部分を決定していました。事前に全く構造がわかっていない新規構造を解析する場合には、タンパク質の特定部位にラベルした重原子による異常散乱を用いて位相を決定し、構造を決定する方法が一般的に用いられます。そこで、本研究では、SACLAにおいてタンパク質微結晶に天然で含まれる硫黄原子によるXFELパルスの異常散乱の検出を試みました。
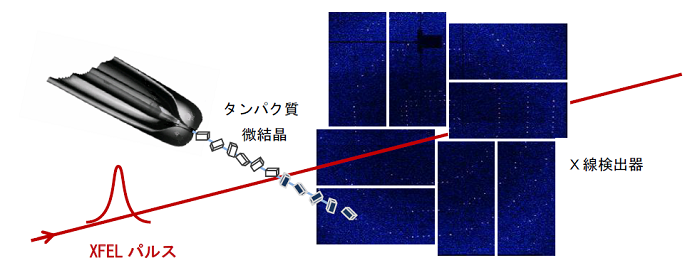
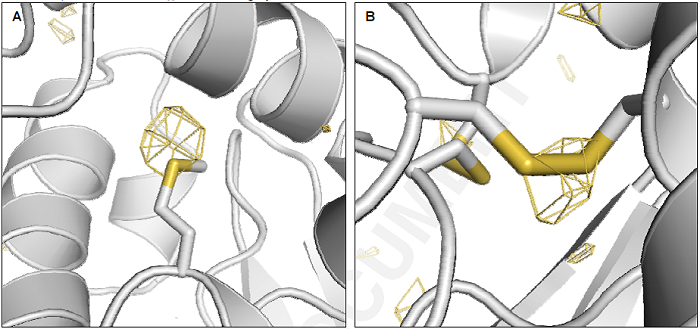
本研究では、構造がすでにわかっているリゾチームタンパク質微結晶にSACLAで得られる強力なX線パルスを照射しました。SACLAの非常に強力なX線パルスを微結晶に照射すると微結晶は損傷を受けます。しかし、SACLAで得られるX線パルスの幅は10 フェムト秒(1秒の百兆分の1)程度と非常に短いため、損傷の影響が微結晶に現れる前のX線回折像を得ることができます。ただし、ここで解決すべき問題がひとつあります。SACLAでは1秒間に数十発のX線パルスが供給されるため、各々のX線パルスが新しい微結晶を照射するように次から次へと微結晶を供給しなければなりません。本研究では、粒径が1ミクロン(1ミクロンは1ミリの1000分の1)程度の微結晶の高濃度白濁液を直径が5ミクロン程度の液体ジェットとして真空中に導入して1ミクロン程度に集光したX線パルスを照射し、X線パルス毎にX線回折像を記録しました(図1)。次から次に(シリアルに)微結晶を導入してXFELの極短X線パルスを用いて微結晶が損傷を受ける前にX線回折像を記録するこの手法はシリアルフェムト秒X線結晶構造解析法と呼ばれています。今回の研究のポイントはシリアルフェムト秒X線結晶構造解析法で記録したX線回折像の各回折斑点の積分強度を精密に評価して、重原子からの異常散乱信号を検出できるかどうかを見極めることでした。2日かけて約600,000枚のX線回折像を撮りましたが、このうち微結晶にXFELが命中できたと思われるものが約420,000枚、このうち各回折斑点の積分強度の解析に使用出来たX線回折像が約44,000枚でした。これをコンピューター上で重ね合わせてX線回折像の回折斑点の積分強度を精密に見積もった結果、タンパク質に天然に含まれている硫黄原子からの異常散乱を検出することに成功しました(図2)。
今回の研究成果は、SACLAによるタンパク質微結晶のシリアルフェムト秒X線構造解析が可能であること、シリアルフェムト秒X線構造解析においてX線回折像の回折斑点の積分強度を評価して特定の原子からの異常散乱信号を評価できることを示したものです。このことによって、これまで構造が決定されていない新規タンパク質についても、SACLAを光源としたタンパク質微結晶のシリアルフェムト秒X線構造解析法に従来から用いられている重原子同型置換法*8、多波長異常散乱法*9といった方法を組み合わせることによって構造を決定する道筋がついたといえます。結晶化が困難なために未だに構造が決定されていないタンパク質の中には、創薬ターゲットとなる膜タンパク質も多く含まれていることから、創薬分野への大きな貢献も、期待されます。
図1.XFELを用いたシリアルフェムト秒X線結晶構造解析実験の概念図
図2.硫黄原子の異常散乱による差異の現れた部位(黄色の部分)
*1 X線自由電子レーザー(XFEL:X-ray Free Electron Laser)
X線領域で発振する自由電子レーザー (Free-Electron Laser) であり、 可干渉性、短いパルス幅、高いピーク輝度を持つ。自由電子レーザーは、 物質中で発光する通常のレーザーと異なり、物質からはぎ取られた自由な電子を加速器の中で光速近くに加速し、 周期的な磁場の中で運動させることにより、レーザー発振を行う。
*2 X線回折
結晶のように周期的な構造を持つ物質に対して、ある波長のX線をいろいろな角度から照射すると、特定の角度では強いX線の反射が起こるが、別の角度では反射がほとんど起こらないという現象を観測できる。これは物質を構成する原子により散乱されたX線が、結晶構造の繰り返しによって強めあったり、打ち消しあったりするためである。この現象をX線回折と呼ぶ。
*3 異常散乱
X線の波長が原子のX線吸収端エネルギーと異なるとき、X線が原子で散乱された時の散乱波の強度は原子散乱因子(実数)の2乗に比例する。しかし、X線吸収端近傍では分散が変化し(これを異常分散と呼ぶ)、原子散乱因子は複素数となり、X線の波長によって原子散乱因子が変化する。この異常分散による散乱因子の異常な振る舞いによる散乱強度変化を異常散乱と呼ぶ。
*4 X線結晶構造解析法
X線回折の結果を解析して結晶内部で原子がどのように配置しているかを決定する手法をX線結晶構造解析と呼ぶ。結晶からのX線回折は個々の原子からの散乱の重ね合わせとなり、結晶の散乱因子(結晶構造因子と呼ぶ)は複素数で記述される。X線の散乱強度は結晶構造因子の絶対値の2乗に比例する。X線結晶構造解析は測定したX線の散乱強度から結晶構造因子を求め、さらにそこから結晶を構成する原子(タンパク質)の立体構造を同定する作業である。
*5 位相
位相とは周期的な現象のひとつの周期中の位置を示す無次元量で、通常は角度で表される。結晶構造因子は結晶によるX線回折現象を表す複素数であり、振幅と位相で記すことができる。X線の散乱強度からは結晶構造因子の絶対値は求められるが、その位相については知ることができない。これを解決しようとする努力を位相問題という。
*6 LCLS
米国スタンフォード線形加速器センター(現在のSLAC国立加速器研究所)で建設された世界で初めてのXFEL施設。Linac Coherent Light Sourceの頭文字をとってLCLSと呼ばれている。2009年12月からから利用運転が開始された。
*7 SACLA
理化学研究所と高輝度光科学研究センターが共同で建設した日本で初めてのXFEL施設。科学技術基本計画における5つの国家基幹技術の1つとして位置付けられ、2006年度から5年間の計画で整備を進めた。2011年3月に施設が完成し、SPring-8 Angstrom Compact free electron LAser の頭文字を取ってSACLAと命名された。2011年6月に最初のX線レーザーを発振、2012年3月から共用運転が開始され、利用実験が始まっている。諸外国と比べて数分の一というコンパクトな施設の規模にも関わらず、 0.1nm以下という世界最短波長のレーザーの生成能力を有する。
*8 重原子同型置換法
構造が未知のタンパク質について、天然型のタンパク質の結晶と水銀原子(Hg)のような重原子を結晶中でタンパク質の特定部位に結合させた複数の重原子同型置換体結晶からの単結晶X線回折により立体構造を解析する手法。X線回折には結晶内の原子の電子密度が寄与するが、重原子の有無による結晶構造因子(振幅)の変化から位相問題を解決する。
*9 多波長異常散乱法
シンクロトロン放射光やXFELのようにX線の波長を複数選択できる場合、重原子、例えばセレン原子(Se)の異常散乱を利用することで位相を決定することが可能である。この方法を多波長異常散乱法と呼ぶ。タンパク質の構造決定では、もっぱら、Se原子の異常散乱を複数の波長で測定し、位相を決定することが行われる。Se原子はタンパク質中にセレノメチオニンとしてメチオニンの代わりに取り込まれる性質があることから、セレノメチオニン置換タンパク質の結晶と取り込まれたSe原子の異常散乱を使った位相決定はタンパク質X線結晶構造解析で定石となっている。

 Home
Home