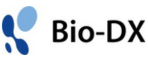本研究成果のポイント
- 3種類のRNAポリメラーゼに共通するテイルがTFIIHの結合部位として機能していることを発見
- 既存の技術では見ることができなかった高い運動性をもつ機能ドメインを含むTFIIHとRNAポリメラーゼIIのドッキング構造のモデル化に成功
- RNAポリメラーゼとTFIIHの相互作用が、転写、転写共役型ヌクレオチド除去修復、細胞増殖に重要であることを解明
- TFIIHがすべてのRNAポリメラーゼの転写やヌクレオチド除去修復に関与していることを証明
概要
横浜市立大学大学院生命医科学研究科の奥田昌彦特任准教授、西村善文特任教授(広島大学大学院統合生命科学研究科長兼任)と東京工業大学生命理工学院の山口雄輝教授の研究グループは、NMR(注1)での構造解析や生化学的手法等を駆使して、RNAポリメラーゼの共通テイルが、転写/修復因子TFIIHに結合し、転写、転写共役型ヌクレオチド除去修復(注2)、細胞増殖に重要であることを明らかにしました。 本研究は、『NucleicAcidsResearch』に掲載されました。また、掲載誌のBreakthrougharticle(注3)に選ばれました。(日本時間7月16日オンライン掲載)
発表内容
【背景】
DNAからRNAを合成する「転写」はRNAポリメラーゼ(RNAP)という酵素が行います。真核生物は3種類のRNAPI、II、IIIを持ち、それぞれリボソーマルRNA、メッセンジャーRNA、トランスファーRNAを合成します。いずれも12個以上のタンパク質(サブユニット)から構成され、そのうち5個は共通しています。
近年の目覚しいクライオ電子顕微鏡(電顕)の技術発展により、RNAPのような複雑な構造が高分解能で解かれるようになりました。大変すばらしい解析法ですが、運動性の高い部分、たとえばRNAPIIが持つ最大サブユニットRPB1のC末にある500残基弱の長いテイルは、一定の構造をとらずふらふらしているため、クライオ電顕構造や結晶構造では見えません。その他のサブユニットはRNAPに共通のサブユニットRPB6を除いてほとんどが見えています。RPB6はN末の41残基、全体の約3分の1もの広い領域が見えていません。RPB6の見えている領域は安定した構造を形成しRNAPIIの内部にあります。そのアミノ酸配列はヒトから酵母までよく保存されています。一方で、見えていない短いN末テイル(NTT)の配列は脊椎動物間でのみ保存されており、脊椎動物の3種類のRNAPは共通した短いテイルを持つことが推測されます。しかし、その普遍性にも関わらず、この短いテイルはこれまで注目されてきませんでした。
【研究成果の内容】
これまで私たちは、基本転写/修復因子TFIIHのp62サブユニットのN末にあるPHドメインと標的タンパク質との複合体の構造をNMRで決定し、認識機構を解明してきました。PHドメインは、様々な転写因子や修復因子と相互作用してTFIIHを機能部位へ先導するため、遺伝子の正常な発現に重要な役割を果たしていると考えられるためです。その研究の過程で、RPB6のNTTの中にPHドメインとの結合部位に似た配列があることに気付きました。ヒトの系で確認したところ、RPB6はNTTでPHドメインと特異的に結合していましたので、RPB6単独およびPHドメインとの複合体の構造をNMRで決定しました。さらに、RPB6単独の構造をRNAPII中のRPB6の構造と比較したところ、興味深い相違が見られました。RPB6は、NTTの一部がβストランド(β1)となって他のβストランドとβシートを形成します。一方で、RNAPII中ではRPB6のβ1がRPB1のβストランドに置き換わっています。このことから、RNAPIIの形成時にRPB6はRPB1とβストランドを交換し、その分長くなったNTTがRNAPIIのテイルになっていることが推測されます。これを踏まえRNAPIIとTFIIHのドッキング構造モデルを構築しました(図1A,1B下)。
RNAP中のRPB6のNTTと同様に、PHドメインもTFIIHのコアから突出し、クライオ電顕構造や結晶構造では見えません。最近私たちは、分子動力学シミュレーションからPHドメインを含んだTFIIHの動的構造モデルを構築しました(参考文献)。そのTFIIHの構造モデルを使いRNAPIIとのドッキング構造を構築しました。転写開始時にはTFIIH、RNAPIIをはじめ様々なタンパク質が集合して転写開始前複合体を形成します(図1B上)。この時、TFIIHのPHドメインは基本転写因子TFIIEαと相互作用します。NTTはそこから離れているので、RNAPIIとTFIIHのドッキングモデル構造が転写開始段階で形成されるとは考えにくいです。開始に続く伸長段階では、開始段階とは違う様々なタンパク(転写伸長因子)がRNAPIIと相互作用しその働きを助けます。活性化された転写伸長複合体の構造が最近他の研究グループによりクライオ電顕で解かれました。転写伸長複合体とTFIIHとのドッキングモデルを構築したところ、転写伸長因子との立体障害はなくTFIIHはRNAPIIと相互作用できました(図1C)。
このRNAPとTFIIHの相互作用が生体内で重要かどうか、PHドメインとの結合に支障をきたす変異や欠損を導入したRPB6変異細胞で調べたところ、増殖が大きく低下しました。
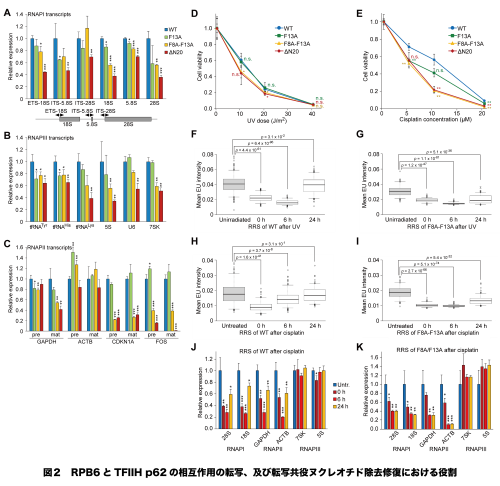
TFIIHがRNAPII以外のRNAPIやRNAPIIIの転写に関与するかは、長い間議論が分かれ結論に至っていません。そこで、PHドメインとRPB6の相互作用変化による各系の転写への影響を調べた結果、RPB6変異細胞ではすべての系で転写が低下しました(図2A—C)。TFIIHは、紫外線照射等で生じたDNAの傷を治すヌクレオチド除去修復にも機能します。この修復には、転写共役型ヌクレオチド除去修復とゲノム(注4)全体で働く経路があります。RPB6変異細胞では、前者の経路のRNAPIとRNAPIIの系で修復の低下がみられました(図2D—K)。以上の結果から、RNAポリメラーゼの共通テイルはTFIIHに結合し転写や修復に重要であること、TFIIHが全RNAPの系に機能することが明らかになりました。
【今後の展開】
今回見出したRNAPとTFIIHとの新しい相互作用が、転写あるいは転写共役型ヌクレオチド除去修復の一連の過程の中でどのように機能するのかを、立体構造に立脚して解明して行きます。また、ヒトの系で見つかったこの相互作用が、酵母など他の生物種で保存され同様な機能を果たすかを明らかにすることも、進化を考える上でとても興味深く重要です。
【研究費】
本研究は、国立研究開発法人日本医療研究開発機構(AMED)「創薬等ライフサイエンス研究支援基盤事業」、文部科学省「先端研究基盤共用促進事業(共用プラットフォーム形成支援プログラム)NMR共用プラットフォーム」の研究の一環で行われました。
【参考資料】
Structural and dynamical insights into the PH domain of p62 in human TFIIH
Masahiko Okuda, Toru Ekimoto, Jun-ichi Kurita, Mitsunori Ikeguchi, Yoshifumi Nishimura
Nucleic Acids Research (2021) 49(5), 2916—2930.
用語解説
(注1)NMR(Nuclear Magnetic Resonance)
強い磁場中で特定の原子核スピンの向きが揃えられた化合物やタンパク質等に対し、ラジオ波を照射して核磁気共鳴させた後、核スピンが元の安定な状態に戻る際に出す信号を観測して、原子の配置などを解析する装置。
(注2)転写共役型ヌクレオチド除去修復
紫外線等、DNAに歪みをもたらすDNA損傷を修復する経路としてヌクレオチド除去修復が存在するが、活発に転写されるDNA領域に生じたDNA損傷は、より深刻なため、速やかに修復される特別な機構があり、転写共役型ヌクレオチド除去修復と呼ばれる。
(注3)Breakthrough Article
独創性、重要性、科学的卓越性の観点から、編集委員会のメンバーとレフリーの推薦に基づき、編集者によって『Nucleic Acids Research』誌の論文の上位2~3%が選ばれる。
(注4)ゲノム
遺伝情報の総体。遺伝子(gene)と染色体(chromosome)を組み合わせた用語で、生物をその生物たらしめるのに必須な最小限の染色体セットと定義されている。
論文情報
- 掲載誌: Nucleic Acids Research
- 論文タイトル: Three human RNA polymerases interact with TFIIH via a common RPB6 subunit
- 著者名: Masahiko Okuda, Tetsufumi Suwa, Hidefumi Suzuki, Yuki Yamaguchi, Yoshifumi Nishimura.
- DOI: https://doi.org/10.1093/nar/gkab612
【お問い合わせ先】
<研究成果に関する窓口>
横浜市立大学 大学院生命医科学研究科 特任教授 西村 善文
Tel:045-508-7211 E-mail:nisimura*yokohama-cu.ac.jp
東京工業大学 生命理工学院 生命理工学系 教授 山口 雄輝
Tel:045-924-5798 E-mail:yyamaguc*@bio.titech.ac.jp
<取材対応に関する窓口>
横浜市立大学 広報課長 上村 一太郎
Tel:045-787-2414 koho*yokohama-cu.ac.jp
広島大学 財務・総務室 広報部 広報グループ
Tel:082-424-3749 koho*office.hiroshima-u.ac.jp
東京工業大学 総務部 広報課
Tel:03-5734-2975 media*jim.titech.ac.jp
(注: *は半角@に置き換えてください)

 Home
Home