脳を持たないとされてきたウニ幼生に、光で行動を調節する「脳のような」の神経細胞群(中枢)を見いだしました。この神経細胞群は、脊椎動物の脳と一部共通する特徴が確認され、後口動物の共通祖先までさかのぼる脳機能の起源に関する新たな示唆を提供する結果となりました。
本研究は、ウニ幼生の前端部神経外胚葉に、非視覚性光感受性ニューロン(「見る」ためではなく、光を感じて応答する神経)の細胞群を同定しました。これにより、脊椎動物の脳に相当する「中枢」が、脳を持たないとされてきた棘皮動物(ウニ)にも存在する可能性が示唆されました。これらの神経細胞群は、光を感知するタンパク質である非視覚オプシン(Opn5L)や、脊椎動物間脳の形成を担うrx、otx、six3、lhx6などの制御遺伝子を発現します。また、この細胞領域を統合的に解析したところ、Opn5Lの機能低下で光依存的な遊泳行動が損なわれることが分かりました。こういった分子特徴は脊椎動物の脳領域のそれと一部重なることから、ウニ幼生に存在する非視覚性の光受容中枢は、後口動物の共通祖先に由来する脳機能の素地を残している可能性を示します。
非視覚オプシンを発現する神経とその周辺領域の発生過程を厳密かつ系統横断的に比較することは、脊椎動物の脳を含む中枢神経の進化や多様化の過程を解く上で、新たな理論や見解を提供すると期待されます。
研究代表者
筑波大学生命環境系
谷口 俊介 准教授
千葉大学大学院医学研究院
露崎 弘毅 特任講師
京都大学大学院理学研究科
山下 高廣 講師
広島大学ゲノム編集イノベーションセンター
山本 卓 教授
研究の背景
進化の過程における中枢神経(脳)の獲得は、ヒトを含む脊索動物の多様化を支えた重要な出来事です。しかし、神経や脳が動物進化のどの段階で生まれ、どの系統でどのように複雑化したのかは、よく分かっていません。脳・中枢神経の主要な役割は外界情報を統合して適応的な運動へ変換することであり、その起源の解明は生物学の根本課題です。
一方、後口動物注1)では、前方神経外胚葉に由来する脳領域(前脳など)が光情報処理の中核を担うと考えられ、表層には、視覚オプシン、深部には非視覚オプシン(いずれも光を感知するタンパク質)が配置されるという脊椎動物との共通性が示唆されています。しかしながら、脊椎動物と最も近縁の棘皮動物門(ウニなど)注2)には集中して存在する「脳領域」が見えにくく、前後軸の明瞭な指標も乏しいため、脊椎動物型の脳がいつ、どのように現れたかをたどることは容易ではありません。
それでも近年、前後軸が明確な成長段階であるウニ幼生で、前脳様の遺伝子発現やそれを結ぶ制御ネットワーク、さらに光などの環境情報を行動へ統合する神経回路が明らかになりつつあります。非視覚オプシン様の受容体も複数同定され、光による遊泳や消化活動の調節に関与する例が報告されています。こうした知見を踏まえ、本研究では、ウニ幼生において非視覚的な光受容を担う「脳様」領域を、遺伝子発現と機能の両面から定義し、脊椎動物の脳に通じる組織化が棘皮動物との共通祖先にさかのぼる可能性を示しました。
研究内容と成果
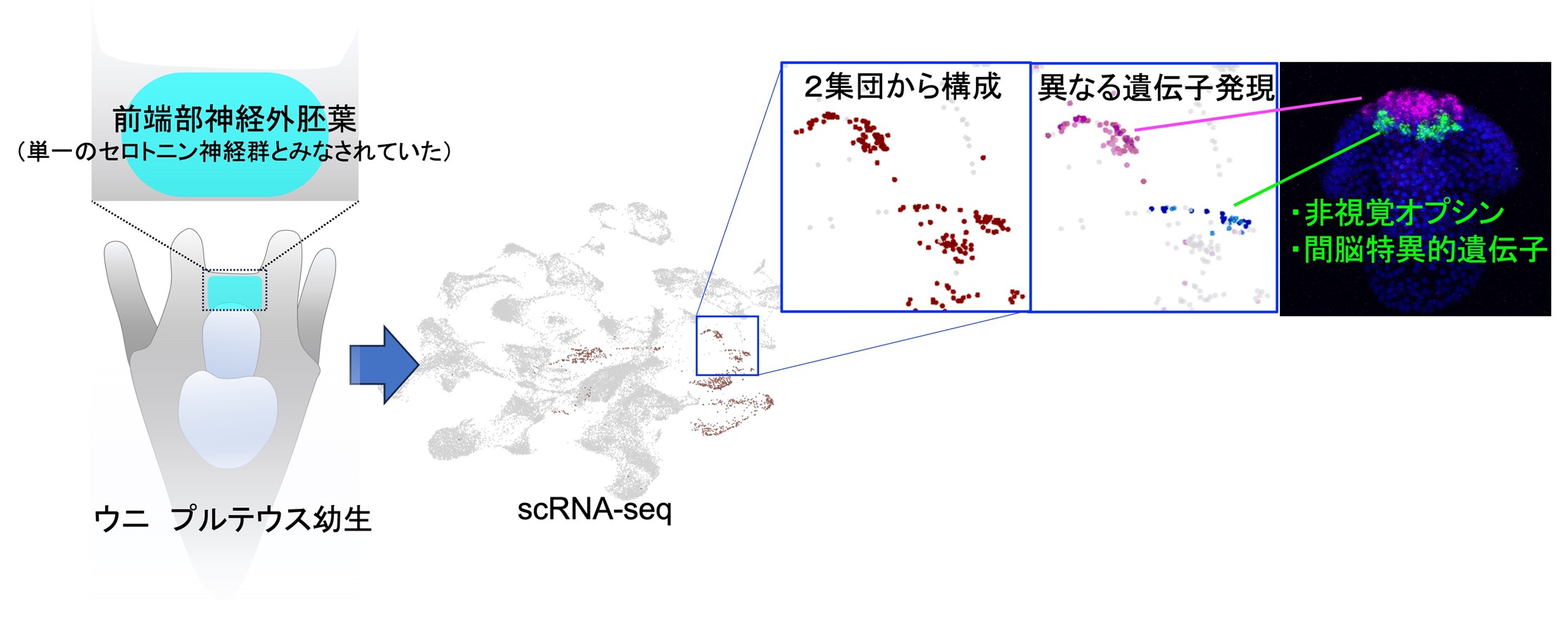
本研究では、バフンウニ(Hemicentrotsu pulcherrimus)幼生の前端部神経外胚葉に存在する「非視覚的な光受容を担う脳様領域(神経細胞群)」を単一細胞RNAシーケンス(scRNA-seq)注3)とin situ ハイブリダイゼーション注4)にて特定し、行動解析によってその機能を統合的に定義しました。幼生期の神経細胞は稀少で検出が難しいため、Delta–Notch による側方抑制注5)を阻害する薬剤を用いて神経細胞数を一時的に増やし、神経集団の解像度を高めました。その結果、これまで一様とみなされがちだったセロトニン注6)作動性ニューロンが、前方群と背側群という二つの集団に分かれており、背側群は非視覚オプシン(Opn5L)を発現していることを見いだしました(図1)。
遺伝子発現の空間解析では、背側群と前方群とでは、形態と配置のいずれもが異なること、さらに特定の転写因子セットが背側群に偏って現れることを明らかにしました。背側群の一部細胞は上皮層から脱落して移動し、前方群の近傍に合流する過程がライブイメージング注7)で観察され、移動性ニューロンとしての性質を持つことが示唆されました。これは、後口動物のうち脊索動物に特有と考えられてきた神経移動が、ウニ幼生にも備わっている可能性を示す重要な知見です。
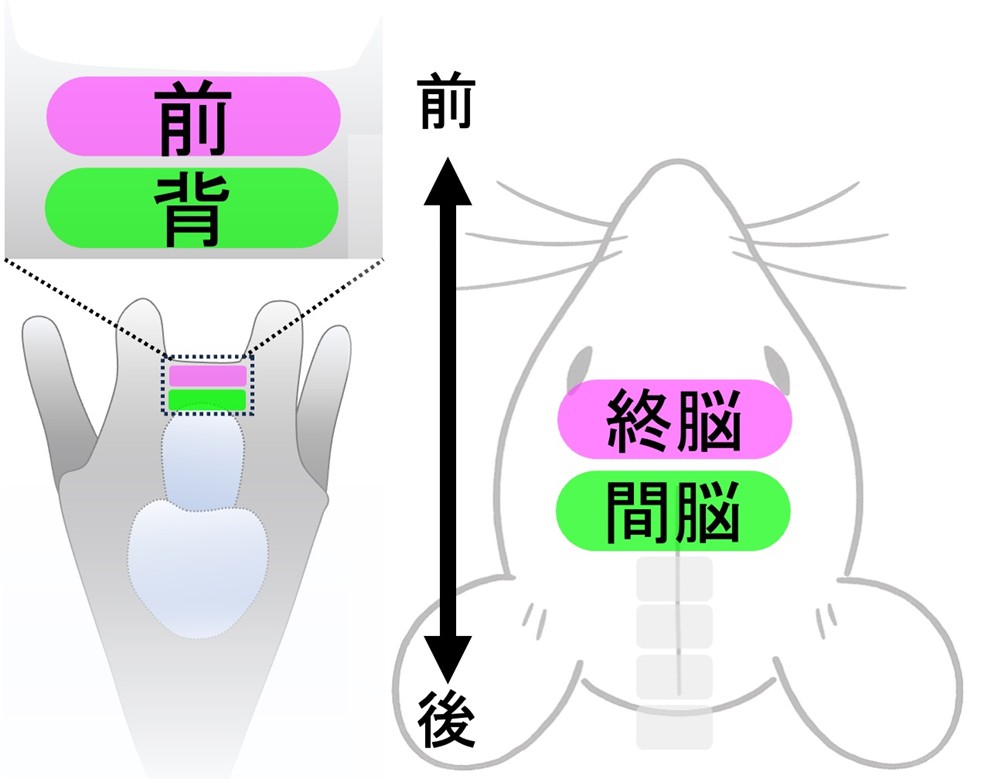
機能面では、Opn5L の機能低下により、連続照明下で沈降(浮遊喪失)行動が有意に抑えられ、光入力が遊泳・浮沈の制御に直接関与することを実証しました。加えて、セロトニン合成関連の制御因子や領域指定因子の発現解析から、当該領域が脊椎動物の終脳/間脳と似た分子設計原理を持つことが示されました(図2)。以上より、ウニ幼生には非視覚光受容を核とする脳様中枢が存在し、環境光情報を行動へ統合する回路の一部が保存されている可能性が高いことが明らかになりました。これは、後口動物共通祖先にさかのぼる脳機能の起点を、具体的な細胞群と遺伝子プログラムとして提示する成果です。
今後の展開
今後は、光→行動の回路に関するより詳細な解析や、比較発生・比較ゲノミクスによる系統横断検証(棘皮・半索・脊索動物での保存/分岐の同定)を進め、非視覚オプシン中枢の普遍性と系統特異性の理解を深めます。最終的には、脳の「はじまり」の設計図を、細胞系譜、遺伝子ネットワーク、行動出力の三層で統合し、脊椎動物脳の起源と多様化に対する新しい指標の提示を目指します。
参考図
図1 (左)ウニ幼生を背側から見た模式図。前端部(水色)領域は前端部神経外胚葉、つまり脊索動物でいうところの脳領域に類似する。しかし、発生初期にここに存在するセロトニン神経は単一のタイプとされ、それほど複雑さはないとみなされていた。(右)この脳領域に対してscRNA-seqを行ったところ、セロトニン神経が2集団から構成され、それぞれ異なる遺伝子発現をしていることが明らかになった。特に体の後方背側に位置する集団は非視覚オプシンとともに、脊索動物では間脳形成に関与している遺伝子群が発現していることが明らかになった。
図2 前後方向に区画分けされたウニ幼生(左図)の脳様領域と、脊椎動物であるマウスの脳(右図)の比較。発現遺伝子プロファイルの比較で、前後軸に沿った類似性が見られた。
用語解説
注1) 後口動物
系統進化上、左右相称動物を大きく二つに分けた時の分類群の一つ。原腸陥入(消化管の元となる原腸が形成される過程)の際の原口(入口部分)が肛門になり、原腸の前端部が体表と接する部分に新たに口ができるグループ。脊索動物と棘皮動物、半索動物を含む。
注2) 棘皮動物門
分類学上の階級(界・門・綱・目・科・属・種)の中で、門で分類した場合のグループの一つ。ウニ、ナマコ、ヒトデ、クモヒトデ、ウミユリの仲間を含む。
注3) 単一細胞RNAシーケンス(scRNA-seq)
多細胞生物の体を構成する細胞それぞれに発現しているmRNAを細胞ごと網羅的に解析する手法。どの細胞にどのような遺伝子が発現しているのかを知ることができる。
注4) in situハイブリダイゼーション
細胞内において特定のmRNAの分布を検出する手法。特定の配列を持つmRNAにラベルをつけ、細胞内のターゲットとなるmRNAと結合(ハイブリダイゼーション)させてラベルを検出する。
注5) Delta-Notchによる側方抑制
隣接する細胞が相互作用することで互いに異なる運命をもたらすメカニズムで、多くの動物の神経細胞分化の過程で見られる現象。DeltaとNotchはいずれも細胞膜上の膜タンパク質で、隣接する一方の細胞のDeltaがもう一方の細胞のNotchと結合することで、この細胞でのDeltaの発現を抑制し、役割の異なる細胞を分化させる。
注6) セロトニン
神経伝達物質の一つ。ヒトの脳にも存在しており、精神安定など、さまざまな機能を果たしている。ウニでは、脳を構成する神経の中で最も早く形成される。
注7)ライブイメージング
光学顕微鏡を用いて生きたままの生き物の動きや細胞の変化を直接観察する手法。今回はウニ胚の細胞膜と核を蛍光タンパク質によって標識し、それをレーザー光で光らせたものを観察した。
研究資金
本研究は、科学技術振興機構(JST) 戦略的創造研究推進事業 さきがけ「多細胞システムにおける細胞間相互作用とそのダイナミクス」研究領域(JPMJPR194C, JPMJPR1945;2019-2022年度)、科学技術振興機構(JST) 研究成果展開事業が助成するA-STEP(JPMJTR204E; 2019-2024年度)、日本学術振興会が助成する科学研究費基盤研究(B)(23K23933; 2022-2025年度)、(C)(23K11312;2023-2027年度)、(若手)(19K20406: 2019-2022 年度)日本医療研究開発機構(AMED) 革新的先端研究開発支援事業 CREST「マルチセンシングネットワークの統合的理解と制御機構の解明による革新的医療技術開発」研究領域(22gm1510007; 2022-2027年度)、東レ科学振興会が助成する東レ科学技術研究助成(2018-2020年度)、武田科学振興財団が助成するライフサイエンス研究奨励(2015年度)による助成によって実施されました。
掲載論文
【題 名】 Non-Visual Photoreceptive Brain Specification in Sea Urchin Larvae
(ウニ幼生における非視覚光受容に関与する脳領域の形成)
【著者名】 #Junko Yaguchi, *#Koki Tsuyuzaki, Ikutaro Sawada, Atsushi Horiuchi, Naoaki Sakamoto, Takashi Yamamoto, Takahiro Yamashita, *Shunsuke Yaguchi (*責任著者、#equal contribution)
【掲載誌】 Nature Communications
【掲載日】 2025年11月19日
【DOI】 10.1038/s41467-025-65628-9
【問い合わせ先】
【研究に関すること】
谷口 俊介(やぐち しゅんすけ)
筑波大学生命環境系/下田臨海実験センター 准教授
TEL: 0558-22-1317
E-mail: yag@shimoda.tsukuba.ac.jp
URL: https://sites.google.com/site/yaguchisea/home
X@urchin_lab
【取材・報道に関すること】
筑波大学 広報室
TEL: 029-853-2040
E-mail: kohositu@un.tsukuba.ac.jp
千葉大学 広報室
TEL: 043-290-2018
E-mail: koho-press@chiba-u.jp
京都大学 広報室 国際広報班
TEL: 075-753-5729
E-mail: comms@mail2.adm.kyoto-u.ac.jp
広島大学 広報室
TEL: 082-424-4518
E-mail: koho@office.hiroshima-u.ac.jp

 Home
Home