このたび、鳥取大学医学部細菌学分野の柴田敏史講師(鳥由来感染症グローバルヘルス研究センター 病態学研究部門)、沖縄科学技術大学院大学(OIST)、広島大学、長崎大学の共同研究グループは、歯周病の原因細菌Porphyromonas gingivalis (ポルフィロモナス・ジンジバリス、以下ジンジバリス菌)がヒトへ感染し歯垢を形成するのに重要な役割を果たす付着器官「Mfa線毛」の立体構造を世界で初めて明らかにしましたのでお知らせいたします。
概要
「ジンジバリス菌」は歯周病だけでなく、誤嚥性肺炎、アルツハイマー病など、様々な病気との関連が指摘されています。しかし、その感染機構の詳細は不明な点も多く、菌を完全に除去する方法はありません。
今回、研究グループは、ジンジバリス菌がヒトの組織や他の細菌に付着する際に利用する「Mfa線毛」のありのままの構造を世界で初めて明らかにしました。また、Mfa線毛がどのように形成されるのかに加え、ヒトの免疫から逃れるために特殊な構造を備えていることや、口腔内のレンサ球菌との結合の仕組みを分子レベルで解明しました。
本研究により、ジンジバリス菌がMfa線毛を使ってヒトの組織へ付着し感染する仕組みが分子レベルで理解できるようになります。今後は、Mfa線毛を標的とした新たな歯周病予防や治療法の開発につながることが期待されます。
なお、本研究成果は2026年6月24日付で国際学術誌「Communications Biology」にてオンライン公開されています。
本研究成果のポイント
- 歯周病の原因細菌ジンジバリス菌が感染に利用する“腕”として機能する付着器官Mfa線毛の立体構造を、クライオ電子顕微鏡※1によって世界で初めて原子レベルで明らかにしました。
- Mfa線毛は細菌が作るプロテアーゼ※2による切断を起点とした「プロテアーゼ依存性ストランド交換機構」によって組み立てられることを明らかにしました。
- Mfa線毛が、ヒトの免疫から逃れるためにカルシウムイオンを利用した特徴的な構造を獲得していることを発見し、ジンジバリス菌の新たな免疫回避機構を明らかにしました。
- Mfa線毛と口腔内病原性レンサ球菌の結合様式をコンピューターシミュレーションで再現し可視化することで、細菌間凝集とバイオフィルム※3(歯垢)形成機構の一端を明らかにしました。
- Mfa線毛の立体構造は歯周病菌の付着や感染を阻害する薬剤開発の標的構造(創薬の鋳型)として利用でき、新たな歯周病予防・治療法の開発につながることが期待されます。
背景
歯周病(歯槽膿漏)は世界で最も蔓延している病気として2001年にギネス記録に認定され、日本でも30歳以上の成人の約8割が罹患、あるいは予備軍であるとされています。
歯周病は歯周ポケットに侵入したジンジバリス菌が組織に付着し、さらに複数の細菌が集まったバイオフィルム(歯垢、デンタルプラーク)を形成することで、歯ぐきに慢性的な炎症を引き起こす感染症です。進行すると歯を支える骨が破壊され歯が抜けてしまいます。さらにジンジバリス菌は口腔内から肺に侵入し誤嚥性肺炎を引き起こしたり、血中へ侵入して全身に広がることで、糖尿病、アルツハイマー、リウマチ、脳梗塞、心疾患、早産など、様々な疾患を発症、悪化させたりします。一方、ジンジバリス菌を完全に排除する方法、薬は未だありません。
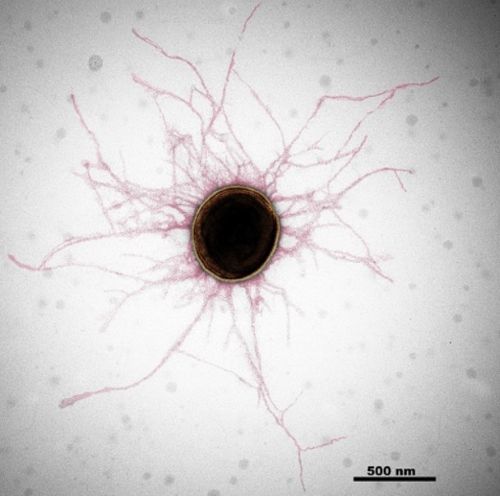
図1:ジンジバリス菌の電子顕微鏡画像
ジンジバリス菌は、線毛と呼ばれる細い繊維状の構造で覆われている(画像処理で赤に彩色)。Mfa線毛とFim線毛を使いヒトに感染する。
病原性細菌が感染するためには、ヒトや動物などの宿主に取り付く必要があります。線毛は細菌が宿主組織や他の細菌と結合するための“腕”の役割を持った繊維状の付着装置です。中でもジンジバリス菌とその近縁種の線毛は、他の細菌線毛とは異なる形成機構で構築されることから5型(V型)線毛として分類されています(参考文献1)。ジンジバリス菌は「Fim線毛」と「Mfa線毛」と呼ばれる2種類のV型線毛を持っており(図1)、それぞれ決まった口腔内のタンパク質や免疫に関わる分子、口腔内細菌と結合します。つまり、2種類の線毛を使い分けてヒトに感染・寄生し病気を引き起こします。
研究グループは以前の研究でFim線毛(第一の腕)の構造と形成機構を明らかにしていました(参考文献2)。今回の研究でMfa線毛(もう一つの腕)の性状を解明したことで、ジンジバリス菌の付着・感染を担う“両腕の全貌”が明らかになりました。
研究成果の内容
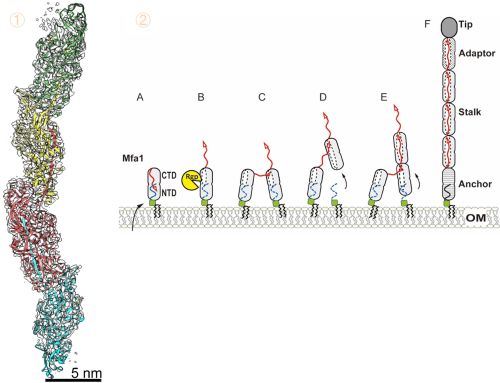
研究グループはMfa線毛を構成するタンパク質Mfa1ピリン※4を試験管内で重合※5させ、クライオ電子顕微鏡による構造解析を行いました。その結果、3.0Å分解能※6でMfa線毛の天然状態の高分解能立体構造を決定することに成功しました(図2①)。この構造情報と生化学的解析結果に基づいてMfa線毛の性状を明らかにしました。
「ストランド交換」による線毛形成機構(図2②)
線毛の部品であるピリンMfa1はジンジバリス菌の表面に輸送されると、N端領域※7がプロテアーゼによって切断されます。すると、内部に折りたたまれていたC端領域のβストランドが大きく構造変化し反転放出されます。その結果、ピリン内部に疎水性の溝が露出します。切断されたピリン同士が近づくと、隣接するピリンのβストランド※8が溝に結合する「ストランド交換」が起こりピリン同士が連結します。この反応が連続的に起こり、Mfa線毛が形成されることが明らかとなりました(図2②A-F)。このような形成機構はFim線毛でも見られることから、V型線毛に共通する普遍的な形成原理であることが確かめられました。
図2:Mfa線毛構造とストランド交換による構築機構モデル
①クライオ電子顕微鏡解析で明らかになったMfa線毛の立体構造。Mfa1ピリン分子が、腕を伸ばすように隣のピリンに結合している。②Mfa線毛構築モデル。Mfa1は菌体の表面に膜タンパク質として輸送されたのち、プロテアーゼRgpBによってNTDの一部が切断される。これをきっかけにC端のストランド(赤矢印)が反転する(A-B)。切断状態のピリン同士が接近するとC端ストランドで結合する(C-D)。この反応が繰り返され線毛は根元伸長し構築される(E)。Mfa1はStalkを構成する、線毛にはその他Tip, Adaptor, Anchorが含まれる(F)。
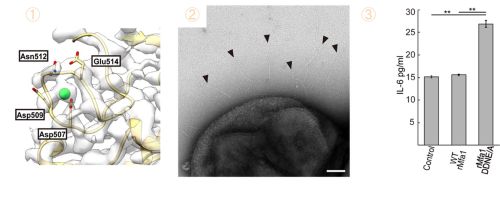
カルシウム結合によるMfa線毛の免疫回避機構
構造解析から、Mfa1分子内部に金属イオンが存在することが判明しました(図3①)。さらにICP-AES(誘導結合プラズマ発光分光分析)による元素分析によって、この金属イオンがカルシウムであることを特定しました。このカルシウムを保持するための4つのアミノ酸を変異させ、カルシウムイオンの結合を妨げても線毛は正常に形成されたので(図3②)、カルシウムイオン結合は線毛の構築に関与していませんでした。一方、この変異型線毛をヒトの培養細胞へ作用させると、免疫応答に関わる炎症性サイトカインIL-6※9の産生を上昇させました(図3③)。これはカルシウムイオン結合領域の構造が変化した変異型Mfa線毛が、ヒトの免疫に認識されやすくなったことを示しています。このことから、ジンジバリス菌は宿主免疫を回避するために進化の過程でMfa線毛にカルシウムイオンを取り込んだと推測できます。
図3:Mfa線毛の免疫回避に関与するカルシウムイオン結合
①Mfa1中のアスパラギン酸(Asp)、アスパラギン(Asn)、グルタミン酸(Glu)を含むループ構造にカルシウムイオン(緑球)が結合している。②カルシウムが結合しない変異型Mfa1も正常に重合し、菌体上に線毛が形成される(矢印)。③カルシウム結合変異型Mfa線毛(rMfa1DDNE/A)は未処理群(Control)、カルシウムイオン結合野生型Mfa線毛(WTrMfa1)と比較して、ヒト培養細胞への免疫誘導性が高くIL-6産生量を増加させた。野生型線毛は免疫を回避しており、未処理群と同等のIL-6産生量であった。
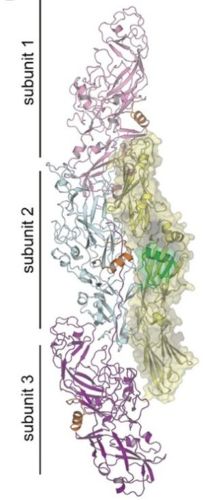
口腔内レンサ球菌との結合様式
Mfa線毛は口腔内のレンサ球菌Streptococcus gordoniiの菌体表面にあるSspBタンパク質と結合し、細菌間共凝集を引き起こします。本研究によって、Mfa線毛表面にあるSspB結合領域の立体構造が明らかになったことで、コンピューターシミュレーションを用いた高精度の分子間相互作用解析が可能となります。これによりMfa線毛とSspBの結合を再現し可視化することができました。
図4: Mfa線毛と口腔内レンサ球菌のSspBとの結合シミュレーション
決定したMfa線毛構造(3分子結合)とすでに解明されているSspB構造(黄色)の結合を、コンピューターシミュレーションで再現した。SspB上でMfa線毛の結合に重要とされる⍺ヘリックスを緑色でハイライトしている。
今後の展開・意義
本成果により、ジンジバリス菌が線毛を介して、どのようにヒトに付着し感染するのか、細菌間で結合しバイオフィルムを形成するのかを分子レベルで理解できるようになります。また、近年の創薬分野研究では、コンピューターを用いて、病気の原因となる標的タンパク質に結合する化合物を仮想的に設計・探索・絞り込む「インシリコ(in silico)創薬」が重要な役割を果たしています。そのためには、標的分子の高分解能で天然状態の立体構造情報が不可欠です。本研究で得られたMfa線毛の詳細な立体構造は、ジンジバリス菌の付着・感染を阻害する分子の設計に向けた創薬標的構造(ドラッグデザインの鋳型)として活用でき、新たな歯周病予防・治療薬開発の基盤となることが期待されます。これらにより歯周病克服を通じた人類の健康維持に大きく貢献することが期待されます。
参考文献
1. Xu, Shoji, Shibata et al. A Distinct Type of Pilus from the Human Microbiome. Cell 165, 690–703 (2016).
2. Shibata et al. Structure of polymerized type V pilin reveals assembly mechanism involving protease-mediated strand exchange. Nat. Microbiol. 5, 830–837 (2020)
用語解説
※1 クライオ電子顕微鏡(構造解析):タンパク質でできた生体分子を急速凍結し、液体窒素の低温下で観察することができる電子顕微鏡。分子のありのまま(天然状態)の構造を高分解能で可視化できます。得られた画像データを画像処理することで分子の立体構造を構築し解析することができます。
※2 プロテアーゼ:タンパク質を切断する酵素。線毛重合にはジンジバリス菌が分泌するジンジパインRgpBと呼ばれるプロテアーゼが関わっています。ジンジパインは歯周組織や血液を分解する病原性因子としての役割もあります。
※3 バイオフィルム:細菌が表面に付着して形成する集合体。細菌は粘着性物質を作り覆うことで、薬剤や免疫に強くなり、除去が困難になります。歯垢(デンタルプラーク)は口腔内バイオフィルムの代表例です。
※4 ピリン:線毛の部品となるタンパク質の総称。
※5 重合:タンパク質がいくつも結合し大きな構造を作ること。Mfa線毛はMfa1ピリンタンパク質が連結して伸長します。
※6 分解能(Å オングストローム):構造解析でどれだけ細かい構造まで見えるかを示す指標。1Åは1億分の1センチメートル。数値が小さいほど高精細構造を示す。3Åの分解能ではタンパク質を構成するアミノ酸が可視化され、金属イオンの存在や分子同士の結合様式を原子レベルに近い精度で理解することができる。
※7 N端、C端(領域):タンパク質はアミノ酸が連結してできており、始まりの部分をN端、終わりの部分をC端と呼びます。
※8 βストランド:タンパク質の骨格構造(二次構造)の一つ。3〜10アミノ酸が連なる紐状構造。
※9 炎症性サイトカインIL-6:免疫細胞などから分泌され、炎症反応を引き起こしたり増強したりする情報伝達タンパク質の一種。細菌やウイルス感染、組織損傷などが起こると産生され、免疫細胞を活性化して体を防御する働きがあります。
論文情報
●題目:Cryo-EM structure of the native assembled Mfa type V pilus from the periodontal pathogen Porphyromonas gingivalis
●著者:Satoshi Shibata*, Hideyuki Matsunami, Kazuhisa Ouhara, Yuri Taniguchi, Makoto Tokoro Schreiber, Alejandro Villar-Brillones, Koji Nakayama, Mikio Shoji and Matthias Wolf* (*責任著者)
●掲載誌:Communications Biology
●DOI:10.1038/s42003-026-10515-2
研究支援
本研究は日本学術振興会(JSPS)科研費 JP19K10083, JP23K06530, JP17K07318, JP20K06581 及び、日本医療研究開発機構(AMED) JP25ama121037 の支援を受けて行いました。
【お問い合わせ先】
<研究に関すること>
鳥取大学医学部医学科感染制御学講座細菌学分野
講師 柴田 敏史
TEL:0859-38-7580
E-mail:sshibata*tottori-u.ac.jp
長崎大学大学院医歯薬学総合研究科 新興感染症病態制御学系専攻
教授 内藤 真理子
TEL:095-819-7648
E-mail:mnaito*nagasaki-u.ac.jp
広島大学大学院医系科学研究科 歯周病態学研究室
講師 應原 一久
TEL:082-257-5663
E-mail:kouhara*hiroshima-u.ac.jp
<取材担当>
鳥取大学米子地区事務部総務課広報係
TEL:0859-38-7037
FAX:0859-38-7029
E-mail: me-kouhou*ml.adm.tottori-u.ac.jp
長崎大学広報戦略本部
TEL:095-819-2007
E-mail:kouhou*ml.nagasaki-u.ac.jp
広島大学総務・広報部広報グループ
TEL:082-424-3701
E-mail:koho*office.hiroshima-u.ac.jp
(*は半角@に置き換えてください)

 Home
Home