(研究に関すること)
広島大学大学院統合生命科学研究科 食品生命科学プログラム
教授 船戸 耕一
Tel:082-424-7923
E-mail:kfunato*hiroshima-u.ac.jp
(報道に関すること)
広島大学 広報室
E-mail:koho*office.hiroshima-u.ac.jp
(*は半角@に置き換えてください)

 Home
Home広島大学大学院統合生命科学研究科の花岡和樹(博士課程後期3年)、松永空也(博士課程前期2年)、清水聡一郎(博士課程前期2年)、生物生産学部の酒井湊士(学部4年)、グラーツ工科大学のHarald Pichler准教授、船戸耕一教授の研究グループは、出芽酵母TDA5がヒトDHRSX遺伝子同じ働きをしていることを発見し、2024年に改訂された3段階のドリコール生合成遠回経路が出芽酵母にも保存されていることを明らかにしました。
この研究では、タンパク質の糖鎖修飾に必要な脂質であるドリコールの生合成に着目して解析を行いました。ドリコールは、小胞体で合成される糖鎖修飾を支える重要な脂質であり、その合成異常はヒトでは先天性糖鎖異常症(CDGs)の原因となります。2024年に、ヒトでDHRSXがドリコール生合成に関与することが発見され、3段階のドリコール生合成遠回経路が新たに提唱されましたが、この遠回経路がヒトで進化的に派生したものなのか、あるいは真核生物に広く保存された仕組みなのかは分かっていませんでした。
本研究グループは、出芽酵母においてTDA5がヒトDHRSXと同じ機能を持つ(機能的オーソログである)ことを見出しました。TDA5を欠損した酵母では、糖鎖修飾の阻害剤であるツニカマイシンに対する感受性が高まり、糖タンパク質CPYの糖鎖修飾異常に加えて、ドリコールの前駆体であるポリプレノールの蓄積とドリコール量の減少が観察されました。これらの異常はヒトDHRSXをTDA5欠損酵母に発現させることで回復したことから、TDA5がDHRSXと同じ機能を担うことが示されました。
今回の研究成果は、2024年にヒトで提唱された3段階のドリコール生合成遠回経路が、出芽酵母にも保存されていることを示唆しています。つまり、遠回経路がヒトに特有のものではなく、真核生物に広く保存された基本的な仕組みであることを示しています。さらに本研究の成果は、ヒトの疾患モデルとしてあるいは薬剤スクリーニングのモデル生物として酵母を活用できる可能性を示しており、ドリコール生合成の異常によって引き起こされる先天性糖鎖異常症(CDGs)の発症機構の理解や治療薬の開発につながることが期待されます。
本研究は、国際科学雑誌『Proceedings of the National Academy of Sciences of the United States of America(PNAS)』に、5月27日に掲載されました。
掲載雑誌名:Proceedings of the National Academy of Sciences of the United States of America(PNAS)
論文名:The revised three-step detour pathway in dolichol biosynthesis is evolutionarily conserved in budding yeast
著者名:Kazuki Hanaoka†, Kuya Matsunaga†, Souichirou Shimizu, Soshi Sakai, Harald Pichler, Kouichi Funato*
(†共同筆頭著者)(*責任著者)
DOI:https://www.pnas.org/doi/10.1073/pnas.2613147123
ドリコールは、ヒト、植物、酵母を含む真核生物の細胞においてタンパク質の糖鎖修飾を支える必須の脂質です。そのため、ドリコール生合成に異常が生じると、酵母では生育不全、ヒトでは先天性糖鎖異常症(CDGs)を引き起こします。
これまで、ドリコールはポリプレノールから1段階の還元反応によって合成されると考えられてきました。このモデルは、ヒトのCDGs原因遺伝子であるSRD5A3(※10)と、その酵母オーソログDFG10(※11)の発見によって強く支持されてきました(Cell, 2010)。ところが2024年に、ヒトのCDGs患者の解析から、DHRSXが新たに発見され、ポリプレノールからポリプレナール(※12)、ポリプレナールからドリカール(※13)、ドリカールからドリコールへと変換される、3段階のドリコール生合成遠回経路が提唱されました(Cell, 2024)。ヒトでは、DHRSXが最初と最後の段階を、SRD5A3がその中間段階の反応を担うと考えられています。
しかし、酵母にはDHRSXに対応する因子がこれまで見つかっておらず、この3段階の遠回経路がヒトで進化的に派生した特別な仕組みなのか、あるいは酵母を含む真核生物に広く保存された基本的な生合成機構なのかは明らかになっていませんでした。
今回、当該研究グループは、ヒトDHRSXと同じ短鎖脱水素酵素(Short-chain Dehydrogenase Reductase: SDR)タンパク質ファミリー(※14)をコードする出芽酵母の遺伝子群に着目し、DHRSXに対応する酵母因子の探索を行いました。その結果、TDA5欠損株が糖鎖修飾の阻害剤であるツニカマイシンに強い感受性を示すこと、糖タンパク質CPYの糖鎖修飾に異常が生じていることを見出しました。これらの結果は、TDA5がドリコール生合成に関与していることを示しています。
次に、TDA5がヒトDHRSXの機能的オーソログであるかを検証しました。そこで、ヒトDHRSXを酵母TDA5欠損株に発現させると、TDA5欠損株のツニカマイシン感受性とCPYの糖鎖修飾異常が回復しました。
また、ポリプレノールとドリコールの量を測定したところ、TDA5欠損株ではポリプレノールが著しく蓄積し、反対にドリコール量は著しく減少しました。これらの表現型もDHRSXの発現によって回復しました(図1)。これらの結果から、TDA5はDHRSXと同様に、ポリプレノールからポリプレナールへの変換、およびドリカールからドリコールへの変換に関わる可能性が強く示されました。
さらに、酵母における既知のドリコール生合成因子DFG10との関係についても解析しました。DFG10はSRD5A3の酵母オーソログとして知られていますが、TDA5とDFG10は互いの欠損を補うことができませんでした。また、TDA5欠損とDFG10欠損を組み合わせた二重欠損株では、単独欠損株よりも強い糖鎖修飾異常が認められました。これらの結果は、TDA5とDFG10が同一の反応経路上で働いているだけでなく、ドリコール生合成経路の中で別の役割を担っている可能性を示しています。
以上のことから、本研究は、ヒトで提唱された3段階のドリコール生合成遠回経路が出芽酵母にも保存されていること明らかにし、真核生物に広く共有された基本的な生物学的機構であることを示した重要な成果であると言えます。
本論文では、DFG10が失われた条件でもドリコールが完全には消失しないことから、酵母には改訂された3段階の遠回経路に加えて、別のバイパス経路が存在する可能性が示唆されました。そのため、この未知のバイパス経路や関与する因子を同定することが今後の重要な課題となります(図2)。
加えて、本研究で得られた酵母での知見は、ヒトの先天性糖鎖異常症(CDGs)をはじめとする関連疾患の発症機構の理解や薬剤開発につながることが期待されます。
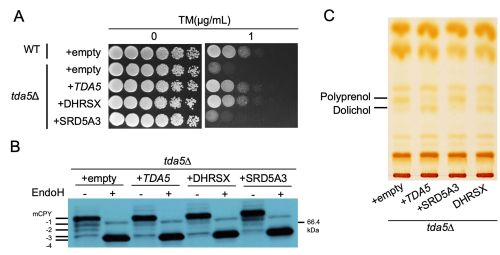
図1.TDA5欠損株におけるツニカマイシン感受性、CPY糖鎖修飾異常、ポリプレノールおよびドリコール異常は、ヒトDHRSXの発現によって回復する
TDA5がヒトDHRSXの機能的オーソログであるかを検証するため、TDA5欠損株(tda5Δ)に酵母TDA5、ヒトDHRSX、ヒトSRD5A3をそれぞれ発現させ、糖鎖修飾阻害剤ツニカマイシン(TM)に対する感受性(A)、CPYの糖鎖修飾異常(B)、ならびにポリプレノールおよびドリコール量(C)を調べました。ツニカマイシン感受性は希釈系列法を用いたスポットアッセイにより評価しました。CPYの糖鎖修飾異常はウエスタンブロット法により解析し、成熟型CPY(mCPY)の下にラダー状に検出される未成熟型バンド(−1から−4)として観察されます。ポリプレノールおよびドリコール量は、薄層クロマトグラフィーを用いた脂質解析により評価しました。TDA5欠損株において、酵母TDA5またはヒトDHRSXを発現させると、ツニカマイシン感受性、CPYの糖鎖修飾異常、ポリプレノールの蓄積、およびドリコール量の低下はいずれも回復しました。
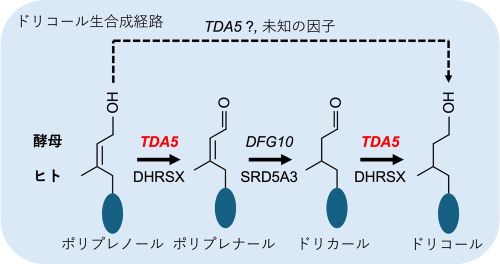
図2.出芽酵母およびヒトにおけるドリコール生合成迂回経路のモデル
ヒトでは、DHRSXとSRD5A3が関与する3段階のドリコール生合成遠回経路が提唱されています(Cell、2024)。本研究は、出芽酵母においてTDA5がDHRSXに対応する遺伝子として機能し、DFG10とともに同様の遠回経路に関与することを示しました。TDA5は、ポリプレノールからポリプレナールへの変換、およびドリカールからドリコールへの変換に関与する可能性があります。また、酵母においては、未知因子によるバイパス経路の存在も示唆されました。
※1 ドリコール
真核生物の細胞に存在する長鎖ポリイソプレノイド脂質。小胞体において、タンパク質に糖鎖を付加する際の糖鎖前駆体の運搬体として働き、糖鎖修飾に必須である。
※2 先天性糖鎖異常症(Congenital Disorders of Glycosylation; CDGs)
糖鎖修飾に関わる遺伝子の異常によって生じる先天性疾患群。糖鎖修飾は多くのタンパク質の機能に必要であるため、その異常は神経系、肝臓、筋肉など全身のさまざまな臓器に影響を及ぼし、発達障害や多臓器症状を引き起こす。ドリコール生合成異常もその原因の一つである。
※3 DHRSX
2024年に、ヒトの先天性糖鎖異常症患者の解析から、ドリコール生合成に関与する因子として見いだされたタンパク質。改訂された3段階のドリコール生合成経路において、最初と最後の反応段階を担うと考えられている。
※4 3段階のドリコール生合成遠回経路
ヒトで提唱された、新しいドリコール生合成経路。従来はポリプレノールからドリコールへの1段階反応が主要経路と考えられていたが、この改訂経路では、ポリプレノールがポリプレナール、ドリカールを経てドリコールへと変換される。
※5 TDA5
出芽酵母に存在する遺伝子。本研究により、ヒトDHRSXの機能的オーソログとして働き、ドリコール生合成に関与することが示された。
※6 オーソログ
異なる生物種の間で、共通の祖先遺伝子に由来し、似た機能を持つ遺伝子のこと。一般に、進化の過程で種が分かれた後も基本的な機能が保存されている。
※7 ツニカマイシン
タンパク質のN結合型糖鎖修飾を阻害する薬剤。ドリコール生合成異常を示す変異株はツニカマイシンに高い感受性を示す。
※8 CPY(Carboxypeptidase Y)
酵母の液胞に存在する糖タンパク質。糖鎖修飾の異常を調べる指標として広く用いられる。
※9 ポリプレノール
3段階のドリコール生合成の出発物質となる長鎖ポリイソプレノイド脂質。3段階のドリコール生合成遠回経路では、ポリプレノール、ポリプレナール、ドリカールを経てドリコールへと変換される。
※10 SRD5A3
先天性糖鎖異常症の原因遺伝子の一つであり、改訂された3段階の迂回経路ではポリプレナールからドリカールへの変換を担うと考えられている。
※11 DFG10
出芽酵母におけるSRD5A3のオーソログ。
※12 ポリプレナール
3段階のドリコール生合成遠回経路における中間体の一つ。ポリプレノールが酸化されて生じる。
※13 ドリカール
3段階のドリコール生合成遠回経路における中間体の一つ。ポリプレナールから生成され、さらに還元されてドリコールとなる。
※14 短鎖脱水素酵素(Short-chain Dehydrogenase Reductase: SDR)ファミリー酸化還元反応を担う酵素群の一つ。生体内でさまざまな脂質や代謝産物の変換に関与する。DHRSXやTDA5はこのファミリーに属する酵素をコードする遺伝子。
(研究に関すること)
広島大学大学院統合生命科学研究科 食品生命科学プログラム
教授 船戸 耕一
Tel:082-424-7923
E-mail:kfunato*hiroshima-u.ac.jp
(報道に関すること)
広島大学 広報室
E-mail:koho*office.hiroshima-u.ac.jp
(*は半角@に置き換えてください)
掲載日 : 2026年06月01日










Copyright © 2003- 広島大学






