(研究に関すること)
広島大学大学院統合生命科学研究科 食品生命科学プログラム
教授 船戸 耕一
Tel:082-424-7923
E-mail:kfunato*hiroshima-u.ac.jp (*を半角の@に変換してください)
(報道に関すること)
広島大学 広報室
E-mail:koho*office.hiroshima-u.ac.jp (*を半角の@に変換してください)

 Home
Home 広島大学大学院統合生命科学研究科の花岡和樹(大学院生)、中里光希(大学院生)、Philipp Schlarmann(外国人特別研究員)、船戸耕一(教授)、酒類総合研究所の金井宗良(主任研究員)、家藤治幸(元部門長)、ジュネーブ大学のHoward Riezman(教授)、セビリア大学のManuel Muñiz(教授)らの国際共同研究グループは、細胞の中でタンパク質が運ばれる仕組みについて、新しい調節メカニズムを明らかにしました。
細胞の中では、新しく作られたタンパク質はまず「小胞体」で形を整えられ、その後「ゴルジ体」へと運ばれて、最終的に必要な場所へ届けられます。この小胞体からゴルジ体への輸送は、COPII小胞と呼ばれる脂質膜で包まれた小さなカプセルのような構造によって行われています。
しかし、細胞がストレスを受けるとタンパク質の形づくりがうまくいかず、不完全なタンパク質が小胞体にたまります。細胞には、こうした異常なタンパク質を修復するまで外に出さない仕組みがありますが、その詳しい仕組みは明らかになっていませんでした。
本研究グループは、酵母を使った研究で、ストレス時に細胞の膜の脂質が変化し、その変化がきっかけとなってタンパク質の輸送が抑えられることを発見しました。特に「YIP3」という遺伝子が重要な役割を担っていることを突き止めました。
YIP3が作るタンパク質Yip3は、輸送用のカプセルを作る際の“足場”となる重要なタンパク質Sec16の働きを抑えます。これによりカプセルが作られなくなり、小胞体からゴルジ体へのタンパク質の移動が低下すると考えられます。
これらの結果から、小胞体はストレスを受けると自ら膜の状態の変化を感知し、タンパク質の流れを調整する仕組みを持っていることが明らかになりました。この仕組みは、神経の病気や糖尿病、がんなどとも関係があると考えられ、本成果は病気の理解や新しい治療法の開発につながる成果として期待されます。
本研究は、国際科学雑誌『Nature Communications』に、令和8年7月3日(日本時間)付で掲載されました。
掲載雑誌名:Nature Communications
論文名:ER sensing of lipid metabolism drives PRA family-dependent regulation of COPII vesicle transport
著者名:Kazuki Hanaoka†, Mitsuki Nakazato†, Philipp Schlarmann†, Hiroki Nakamura, Mei Kato, Ryoko Ikema, Mizuki Iguchi, Katsuki Eto, Takefumi Karashima, Atsuko Ikeda, Yukari Yabuki, Javier Manzano-Lopez, Auxiliadora Aguilera-Romero, Susana Sabido-Bozo, Ana Maria Perez-Linero, Muneyoshi Kanai, Haruyuki Iefuji, Isabelle Riezman, Howard Riezman, Manuel Muñiz, Kouichi Funato*
(†共同筆頭著者)(*責任著者)
DOI:https://doi.org/10.1038/s41467-026-75057-x
私たちの体をつくる細胞では、タンパク質や脂質が正しい場所へ運ばれることで、生命活動が維持されています。なかでも小胞体は、タンパク質や脂質が作られる重要な細胞小器官(※9)です。小胞体で作られたタンパク質や脂質は、COPII小胞と呼ばれる輸送小胞に包まれてゴルジ体へ運ばれ、その後それぞれの目的地へ届けられます。この小胞体からゴルジ体への輸送は、細胞内物流の出発点にあたり、酵母からヒトまで真核生物に広く保存された基本的な仕組みです。
COPII小胞は、小胞体膜上のER exit site(ERES)(※10)と呼ばれる特定の領域で形成されます。ERESでは、COPII小胞形成に必要なタンパク質群が集まり、Sec16と呼ばれるタンパク質がその集合を支える足場として働くことで、小胞形成が起こります。つまり、ERESは小胞体からの輸送を開始するための“出荷口”であり、Sec16はその出荷口を作るための“土台”として重要な役割を担っています。
一方で、小胞体は常に一定の状態に保たれているわけではありません。細胞がストレスにさらされると、タンパク質の正常な折りたたみが妨げられ、異常なタンパク質が生じて小胞体に蓄積します。この蓄積は、異常タンパク質が適切に修復されるまで他の場所へ移動するのを防ぐ、いわゆる小胞体の品質管理機構によるものと考えられています。しかし、小胞体ストレスがどのようにしてCOPII小胞の形成抑制へと伝達されるのかについては、これまで十分に解明されていませんでした。
本研究グループは、モデル生物である出芽酵母を用いて、小胞体ストレス時にCOPII小胞によるタンパク質輸送がどのように調節されるのかを解析しました。
まず、小胞体ストレスを誘導した細胞では、リン脂質の一種であるホスファチジン酸(PA)の量が増加することを見出しました(図1A)。PAは、小胞体膜上で転写制御因子Opi1の局在を制御することで、転写因子Ino2/Ino4の働きを調節する脂質として知られています。そこで本研究グループは、PA–Opi1–Ino2/Ino4転写制御系(※11)の下流で、COPII小胞輸送を抑制する因子の探索を行いました。
広範な遺伝学的解析の結果、Ino2/Ino4の下流で発現が制御され、COPII小胞輸送を調節する新たな因子として、YIP3遺伝子を同定しました。YIP3がコードするYip3タンパク質は、COPII小胞輸送を抑制する因子として働くことが分かりました。実際に、小胞体ストレスを誘導すると、Yip3タンパク質の量が増加することが観察されました(図1B)。
次に、Yip3がどのようにCOPII小胞形成を抑制するのかを調べました。COPII小胞は、小胞体膜上のERESで形成されます。ERESでは、Sec16がCOPII小胞形成に必要なタンパク質群を集める“土台”として働きます。本研究では、蛍光顕微鏡観察によりSec16が小胞体膜上にどの程度集まるかを調べました。その結果、小胞体ストレス時にはSec16のERESへの集積が低下することが分かりました(図1C)。
以上の結果から、小胞体ストレス時には、小胞体膜におけるPA量が増加し、Opi1–Ino2/Ino4転写制御系を介してYIP3遺伝子の発現が促進し、増加したYip3タンパク質がERESにおけるSec16の集積を抑制することで、COPII小胞の形成を阻害することが明らかになりました(図2)。
本研究は、小胞体が自身の膜脂質の状態を手がかりとして、転写を介して細胞内物流の出荷量を調節する仕組みを備えていることを示したものです。
本研究により、小胞体ストレス時に小胞体膜のホスファチジン酸(PA)レベルが増加し、その情報が転写を介してCOPII小胞の形成の抑制につながることが明らかになりました。
一方で、本研究では、Yip3とSec16は細胞内での局在が完全には一致せず、両者の明確な物理的相互作用も確認されませんでした。このことから、Yip3はSec16を直接制御しているのではなく、別の因子を介してSec16のERESへの集積を制御している可能性があります。今後は、Yip3がどの分子を介してSec16の集積を抑制するのかを明らかにすることが重要です。
また、Opi1–Ino2/Ino4転写制御系の下流には、Yip3以外にもCOPII小胞形成の最初のステップを制御する未知因子(XXX)が存在する可能性が示されたことから(図2)、これらを同定することにより、小胞体ストレス時の輸送制御機構の全容解明に大きく貢献すると考えられます。
Yip3はPRAファミリー(※12)に属するタンパク質であり、このファミリーは酵母からヒトまで広く保存されています。そのため、今回明らかになった仕組みは、ヒトを含む高等真核生物における小胞体ストレス時のタンパク質輸送制御メカニズムを理解する上でも重要な手がかりになります。
小胞体ストレスは、神経変性疾患、糖尿病、がんをはじめとする多様な疾患の病態に関与しています。本成果を基軸として、脂質とPRAファミリータンパク質を介したCOPII小胞の形成制御メカニズムおよび小胞体ストレス応答のメカニズムの解明が進展すれば、分泌不全、小胞体恒常性の破綻、あるいは脂質代謝異常を基盤とする諸疾患の病因・病態の理解に大きく貢献し、新規治療法の開発等につながることが期待されます。
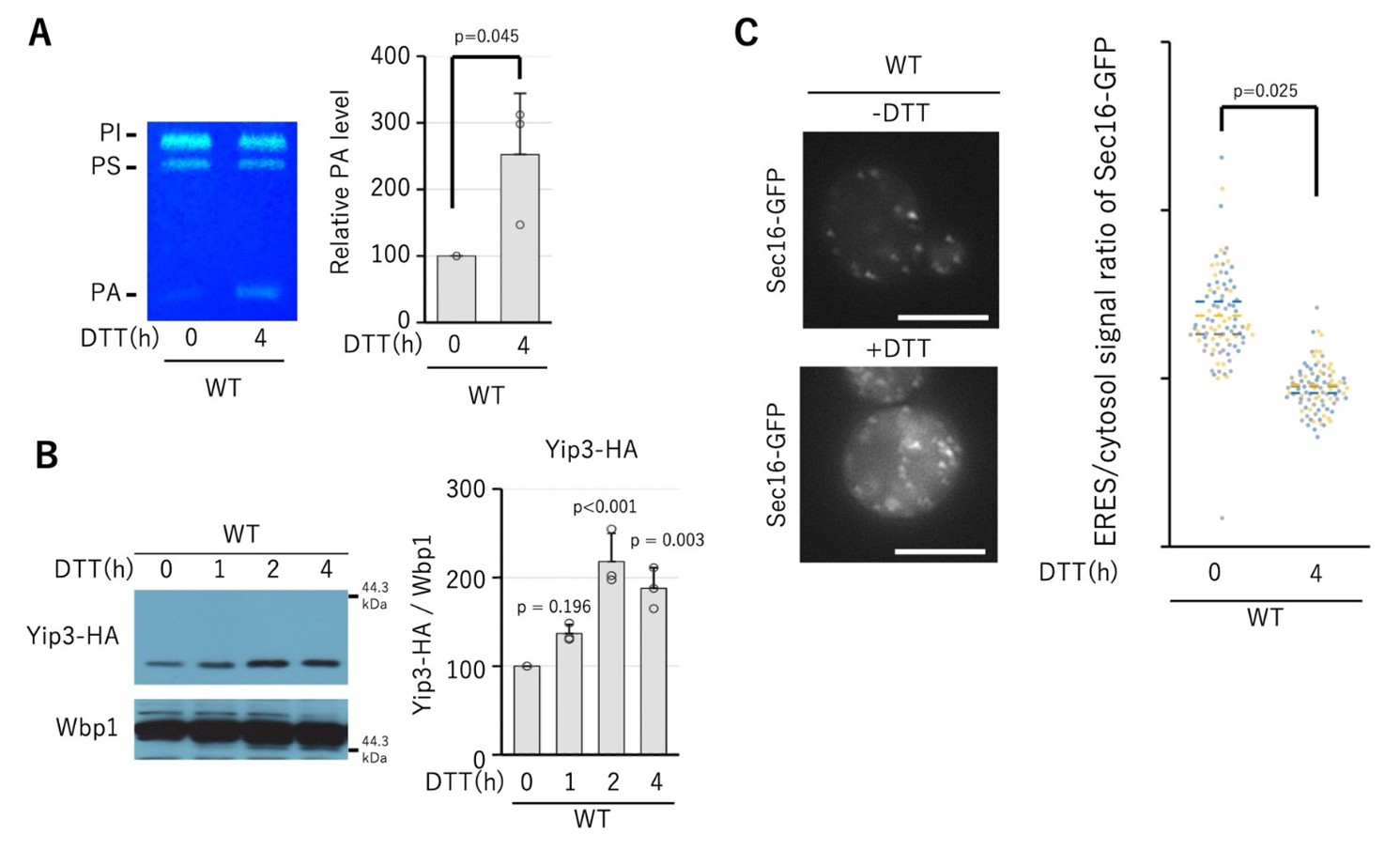
図1.小胞体ストレスは、ホスファチジン酸(PA)量とYip3量を増加させ、Sec16のERESへの集積を低下させる
A 野生株に、小胞体ストレスを引き起こす還元剤DTT(ジチオスレイトール)を4時間処理し、細胞内のPA量を薄層クロマトグラフィー(TLC)により解析した。その結果、DTT処理によりPA量が増加することが確認された。
B YIP3遺伝子にHAタグを付加したYip3-HAを発現する細胞にDTTを1、2、4時間処理し、ウエスタンブロット法によりYip3-HA量を解析した。その結果、小胞体ストレスによりYip3-HA量が時間依存的に増加することが確認された。
C Sec16にGFPタグを付加したSec16-GFPを発現する細胞にDTTを4時間処理し、蛍光顕微鏡によりSec16-GFPの細胞内局在を観察した。DTT未処理の細胞では、Sec16-GFPは小胞体膜上にドット状(ER exit site)に観察され、ドットと細胞質のシグナル強度比は高い値を示した。一方、DTT処理を行った細胞では、Sec16-GFPのドット状シグナルが減少し、細胞質に広く拡散したシグナルが観察され、ドットと細胞質のシグナル強度比は低下した。この結果は、小胞体ストレスにより、COPII小胞形成の“土台”となるSec16がERESに集まり難くなることを示している。
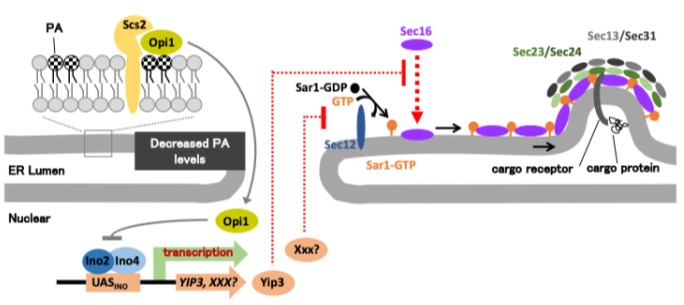
図2.小胞体膜のホスファチジン酸(PA)レベルを感知し、COPII小胞の形成を抑制する仕組みのモデル
小胞体膜のPAレベルが低い場合、Opi1は核内へ移行し、Ino2/Ino4による転写活性を抑制する。その結果、YIP3遺伝子や未知因子XXXの発現が低下し、COPII小胞の形成の抑制が解除される。
一方、小胞体ストレスにより小胞体膜のPAレベルが上昇すると、Opi1は小胞体膜上に留まり、Ino2/Ino4による転写活性化が維持される。その結果、YIP3遺伝子の発現が高まり、Yip3タンパク質がCOPII小胞の形成の足場タンパク質Sec16のERESへの集積を妨げる。これにより、COPII小胞を作るための“土台”が構築されず、小胞体からゴルジ体へのタンパク質輸送が制限される。
本研究は、Yip3以外にも、COPII小胞形成の最初のステップ(Sar1の小胞体膜への結合)を制御する未知因子XXXがIno2/Ino4の下流に存在する可能性を提示しました。これらの結果から、小胞体はPAレベルの変化を手がかりとして、転写を介してCOPII小胞形成を複数の段階で調節していると考えられる。
※1 小胞体
真核生物の細胞内に存在する細胞小器官の一つ。タンパク質や脂質が作られる重要な場所であり、特に分泌タンパク質や膜タンパク質は小胞体で正常に折りたたまれた後、ゴルジ体へ輸送される。
※2 COPII小胞
小胞体からゴルジ体へタンパク質や脂質を運ぶ輸送小胞。小胞体膜の一部がくびれ切れて形成される脂質膜で包まれた小さな輸送カプセルであり、酵母からヒトまで広く保存された細胞内輸送小胞として機能する。
※3 ゴルジ体
小胞体から運ばれてきたタンパク質や脂質をさらに修飾し、細胞内外の適切な場所へ仕分ける細胞小器官。細胞内物流における集荷・仕分けセンターのような役割を持つ。
※4 出芽酵母
パンやお酒の発酵にも利用される単細胞の真核生物。ヒトと共通する多くの基本的な生命現象と仕組みを持ち、遺伝子操作や細胞内の観察が比較的容易であることから、生命科学研究のモデル生物として広く用いられている。
※5 小胞体ストレス
小胞体内でタンパク質の折りたたみがうまく進まず、異常なタンパク質が蓄積することで生じる細胞内ストレス。小胞体ストレスは神経変性疾患、糖尿病、がんなど、さまざまな疾患との関連が知られている。
※6 ホスファチジン酸(PA)
細胞膜や細胞小器官の膜を構成するリン脂質の一種。小さな親水性の頭部と脂肪酸からなる疎水性の尾部を持ち、膜の形や曲がりやすさに影響を与える特徴がある。膜脂質の材料となるだけでなく、細胞内の情報伝達や脂質代謝の調節にも関わる。本研究では、小胞体ストレス時にPA量が増加し、COPII小胞の形成を抑制することが示された。
※7 YIP3遺伝子
出芽酵母に存在する遺伝子で、PRAファミリーに属するYip3タンパク質をコードする。本研究により、YIP3はOpi1–Ino2/Ino4転写制御系の下流で発現が制御され、Sec16のERESへの集積を抑えることでCOPII小胞の形成を抑制する新たな因子として同定された。
※8 Sec16
COPII小胞形成に必要な足場タンパク質。小胞体膜上のERESに集まり、COPII小胞を作るために必要なタンパク質群を集める“土台”のような役割を担う。
※9 細胞小器官
細胞の中で特定の機能を担う構造体。多くは脂質膜で区切られた小さな区画として存在し、小胞体、ゴルジ体、ミトコンドリア、液胞、核などが含まれる。それぞれが役割を分担することで、細胞の生命活動が維持されている。
※10 ER exit site(ERES)
小胞体膜上に存在する、COPII小胞が形成される特定の領域。小胞体からゴルジ体への輸送が始まる“出荷口”にあたり、Sec16などのタンパク質が集まってCOPII小胞の形成を支える。
※11 Opi1–Ino2/Ino4転写制御系
酵母において、細胞内の代謝状態に応じて遺伝子発現を調節する転写制御システム。Ino2/Ino4は遺伝子発現を促進する転写因子として働き、Opi1はその働きを抑える転写制御因子として機能する。Opi1は小胞体膜上のPA量に応じて局在を変えるため、細胞はこの仕組みを介して小胞体膜のPA量の変化を感知する。本研究では、この転写制御系がYIP3遺伝子の発現を制御し、COPII小胞の形成の調節につながることが示された。
※12 PRAファミリー
酵母からヒトまで広く保存されている膜タンパク質ファミリー。PRAはPrenylated Rab Acceptorに由来する名称で、細胞内輸送に関わるタンパク質群として知られている。Yip3は出芽酵母のPRAファミリータンパク質であり、本研究ではCOPII小胞の形成を抑制する因子として同定された。
(研究に関すること)
広島大学大学院統合生命科学研究科 食品生命科学プログラム
教授 船戸 耕一
Tel:082-424-7923
E-mail:kfunato*hiroshima-u.ac.jp (*を半角の@に変換してください)
(報道に関すること)
広島大学 広報室
E-mail:koho*office.hiroshima-u.ac.jp (*を半角の@に変換してください)
掲載日 : 2026年07月09日










Copyright © 2003- 広島大学






